
IF ASKED, most people would probably say that the purpose of sex is to reproduce. This is not correct. Many species reproduce without sex. For example, there are geckoes in South Pacific islands who reproduce asexually. Many other species do too, including whiptail lizards from the American southwest. In these species, the eggs carry the full complement of genes to begin with. (They are formed through mitosis, not meiosis.) Hence, the eggs in such species do not need to be fertilized to develop into embryos. Because the eggs do not need fertilization, males do not exist in these species, which consist solely of females. Thus reproduction is perfectly possible without sex, and the purpose of sex is to provide the offspring with a mixture of genes from two parents. Gene mixing is the purpose of sex, not reproduction per se.
What about the role of “male” and “female” in this equation, and must it be a binary system? When asked, most people define male and female on the basis of external appearance. A male lion has his mane, a rooster his cockle-doodle-do, and so forth. A closer inspection reveals different genitals—in mammals, males with a penis, and females with a vagina. A look inside the body reveals males with testes and females with ovaries. In mammals, a still deeper look into the cells reveals an XY set of sex chromosomes in males and an XX set in females.
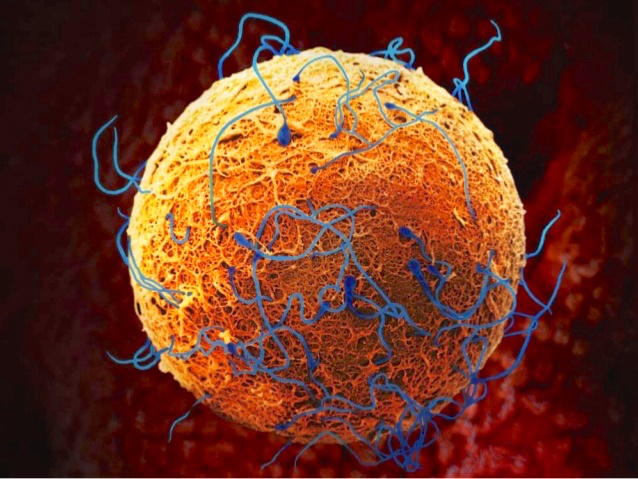 You may be surprised to learn, then, that these traits do not define male and female for biologists. These traits vary too much among individuals within a species and especially across species to serve as defining traits. Instead, to have a definition of male and female that applies to all living things, from seaweed to sea lions, from mice to men, biologists define a male as an individual who produces only sperm and a female as one who produces only eggs throughout their lives, while a hermaphrodite is an individual who produces both sperm and eggs over the life cycle.
You may be surprised to learn, then, that these traits do not define male and female for biologists. These traits vary too much among individuals within a species and especially across species to serve as defining traits. Instead, to have a definition of male and female that applies to all living things, from seaweed to sea lions, from mice to men, biologists define a male as an individual who produces only sperm and a female as one who produces only eggs throughout their lives, while a hermaphrodite is an individual who produces both sperm and eggs over the life cycle.
The question then becomes, what is a sperm and what is an egg? Each gamete is a cell that contains half of the genetic material of an adult. Two gametes fuse to produce the cell that multiplies to become an embryo. The gametes are of very unequal size, and there are only two sizes per species. This is the only binary that exists. The individuals in virtually all sexually reproducing species produce only two sizes of gametes, one large and one small. There are almost no species whose members produce sizes varying continuously from small to large. Moreover, if the individuals of a species produce gametes of only one size, such as some fungi, then male and female are not defined for that species. So, practically speaking, the only universal sex binary in biology is the dichotomy between egg and sperm.
The prevalence of hermaphroditic species among both plants and animals can shed light on male and female sex roles in many other species. For starters, there are three types of hermaphrodite. A simultaneous hermaphrodite is an individual who produces both sperm and eggs at the same time. A sequential hermaphrodite produces eggs and sperm at different times during life, and these come in two varieties: protandrous (male first, then female) and protogynous (the reverse). Sequential hermaphrodites change sex during life, shifting from making sperm to making eggs, or vice versa. A sequential hermaphrodite also changes in incidental characteristics, such as sex-specific body colors, shapes, and behavior.
Most plants are hermaphroditic. For example, a flower typically contains anthers that disperse pollen and a stigma with a sticky surface on which pollen grains are deposited by a pollinator or by the wind. The anthers are the male part of the plant, the pollen grains are the sperm, and the stigma and the ovule at the base of the stigma that becomes the seed comprise the female part. Hermaphrodites are the rule, not the exception, throughout the plant kingdom. Only about six percent of plant species have separate sexes.
Animals, too, present an enormous number of hermaphroditic species, including many marine invertebrates, such as sea urchins, snails, barnacles, and corals. Even among vertebrates, many species are hermaphroditic. If one goes snorkeling on a coral reef, about fifty percent of the fish one sees belong to hermaphroditic species: parrot fish, wrasses, groupers, etc. Across both marine and terrestrial environments, about six percent of all animal species are hermaphroditic. However, if insects are not counted, the percentage rises to about 33 percent of animal species. Although hermaphroditic animal species are common in the marine environment, they are relatively rare in terrestrial environments, an unexplained fact that has invited fun but unproven conjectures over the years.
Sex Roles
Popular discourse about sex roles envisions the male as capable of fertilizing many females with an almost infinite supply of sperm, while the female is limited to producing a finite number of large eggs. Hence, females are in control; they make the choice of a mate lest they waste their expensive eggs on inferior goods. Meanwhile, males are supposed to enjoy the promiscuous pursuit of as many conquests as possible.
And yet, in most fish species, parental care of eggs is provided by males. (In mammals, the care is primarily by females, and in birds by both sexes.) For example, in seahorses, the males have a skin flap on their bellies into which females deposit their eggs, causing the male to become, so to speak, pregnant. As a result, females in some seahorse species can produce eggs faster than the males can give birth to those incubating. Hence the females can mate with many more males than the reverse, turning the traditional balance of supply and demand on its head. Sex-role reversed species such as the seahorse demonstrate that there is no necessary connection between gamete size and sex role.
 Another popular belief is that males are naturally larger than females, offering protection but also exerting control. Deep sea angler fish offer an interesting counter-example. An angler fish is about the size of golf ball and so named because of the bait-like tassel at the end of a front-facing dorsal spine. The angler is a fisherman—actually, a fisherwoman. The large fish depicted here is a female. The small bumps on its bottom are males, sometimes called dwarf males or parasitic males. They are physically attached to the female, in some species even to the point of being connected with the female’s blood circulatory system. Two males are attached to this female, a mating arrangement called polyandry.
Another popular belief is that males are naturally larger than females, offering protection but also exerting control. Deep sea angler fish offer an interesting counter-example. An angler fish is about the size of golf ball and so named because of the bait-like tassel at the end of a front-facing dorsal spine. The angler is a fisherman—actually, a fisherwoman. The large fish depicted here is a female. The small bumps on its bottom are males, sometimes called dwarf males or parasitic males. They are physically attached to the female, in some species even to the point of being connected with the female’s blood circulatory system. Two males are attached to this female, a mating arrangement called polyandry.
It is often said that when males provide no easily discernible paternal care, then all they bring to their matings are their genes, with the work of raising the young defaulting to the female. This is not correct. If it were, all males would be like angler fish, no more than heat-seeking ballistic testes. The fact that males are generally whole organisms themselves rather than convenient accessories carried by the females suggests that males bring more to the table than solely their genes—even if it is not always obvious just what they bring.
Gender
“Gender,” as distinct from sex, is normally thought of as pertaining uniquely to humans, something that’s constructed by culture and finding highly divergent expression in different cultures. What could gender mean when applied to animals? For animals, I take gender to mean the appearance, behavior, and life history of a sexed body, which is a body classified according to the size of the gametes produced. Thus, gender is appearance plus action—how an organism uses its morphology (color and shape) and behavior to carry out a reproductive role.
As we have seen, sex at the whole organism level is not a binary, even though at the gamete level the egg/sperm size distinction does constitute a binary. In fact, gender is even less binary than sex. Even in species with exactly two sexes classified as male and female, the number of genders can be greater than two. There may be, say, two genders of males, as when those with the smaller gamete come in two distinct types based on their appearance, behavior, and life trajectory.
Sandpipers, for example, are a species with three male genders and one female gender. Ruffs are sandpipers, shorebirds that breed in summer in northern Europe. Ruffs owe their name to a ring of feathers that males have around their necks. Shown here is the male gender with a dark ruff, a white ruff, and no ruff. Below is the female, also with no ruff. These genders are genetic, and about twenty percent of the males with a ruff have the white variety and eighty percent the black variety.

Ruffs often mate in leks, which are in effect male “red-light districts” where males congregate to attract females. The black-ruffed males defend small “courts” within the lek and within these each displays to the visiting females. The white-ruffed males do not defend courts within the lek and instead keep company with the females as they feed. What’s remarkable is the interaction between the black- and white-ruffed males. When a white-ruffed male is nearby and a black-ruffed male is alone on a court, the black-ruffed male does a little dance that invites the white-ruffed male to join him on the court he has been defending. Females who then arrive at the lek to mate prefer a black-white team of males over a lone black-ruffed male. Both males jointly court and then mate with the female. A black-ruffed male obtains more matings when a white-ruffed male is present than when by himself, even though the matings are shared. It’s not clear why a female finds a team more attractive than a single black-ruffed male. Most investigators seem to assume that a female automatically finds two males better than one—the more masculinity the better. If more total masculinity is so important, then two black-ruffed males could simply team up with each other. Why two genders?
My hunch has been that a white-ruffed male builds relationships with females while he is with them away from the lek. While the black-ruffed male is defending a court against other black-ruffed males, the white-ruffed male is flying with females in the field and presumably getting to know them. Perhaps the white-ruffed male can, so to speak, make introductions when the females arrive at the lek. He can act as a go-between, a marriage broker, who can introduce a black-ruffed male to a female and vouch for his safety. Whatever the explanation, this example serves to illustrate the phenomenon of gender multiplicity in animals. Two sexes does not imply only two genders.
Finally, a black-ruffed male has also been observed mating with a ruff-less male. This is an example in the wild of a homosexual mating—i.e., between two individuals who produce the same size of gamete. However, this example is also heterogenderal because the two individuals belong to different male genders.
Even in species that have only two sexes and two genders, the gender categories are not absolute, and transgender individuals occur. The best studied example of this phenomenon occurs in a set of sun angel hummingbird species from the Andes. Male sun angel hummingbirds have colorful feathers on their throats called a gorget. A female with a gorget is referred to here as a “masculine female.” She also has a comparatively shorter bill. Conversely, “feminine males” also exist with special female traits such as a longer bill.
Of 42 hummingbird species surveyed throughout the Andes, seven had both masculine females and feminine males, nine had masculine females and no feminine males, two had feminine males and no masculine females, and 24 had neither masculine females nor feminine males. Pooling the species with either masculine females, feminine males, or both, revealed that 52 percent of the females were masculine and two percent of the males were feminine.
Males use their gorgets in territorial defense of the common short flowers that fit their shorter bills. Masculine females can, like the males, defend a territory of short flowers to feed in. Conversely, the feminine males have longer bills than masculine males, even longer than feminine females. Hence, feminine males use different flowers from the masculine males, namely relatively rare long tubular flowers that do not need to be defended in a territory. Thus, the masculine females occupy slightly different niches, i.e., they have slightly different occupations, from the feminine females. Conversely feminine males also occupy somewhat different niches from the masculine males. We see then that gender expression in birds in part reflects a gendered difference in occupation, and transgender birds are those whose occupation crosses over into the occupation typical of the other gender.
Homosexuality
By “homosexuality,” I refer to same-sex mounting and to mutual touching of genitals. Today, the reality of extensive homosexuality among animals is generally accepted among biologists. The 300 or so known examples of homosexuality in animals, surely an underestimate, reveal much diversity. In some species only males are homosexual, in some, only females, and in others, both sexes. In almost all, the homosexuality is mixed in with heterosexuality by the same individual. In some species, a small fraction participates in homosexuality; in others, like our closest relative the bonobo chimpanzee, every animal does. But rather than wondering why homosexuality exists in some animals, we may well ask why homosexual behavior is not dominant, perhaps mixed in with some minimal amount of heterosexuality to ensure reproduction?
Homosexuality is widely distributed across many higher taxa and is especially well-documented in birds, mammals, and insects. This wide distribution implies that homosexuality has originated many times in the animal kingdom. Concerning primates, their family tree indicates a pattern. From its base near the ground, the primate tree trunk splits first into the prosimians and the anthropoids. The prosimian branch, including bush babies, lemurs, and tarsiers, appear to have only incidental same-sex mounting and no same-sex courtship. The anthropoid branch splits into two sub-branches, New World and Old World primates. The former, which includes marmosets, tamarins, and monkeys with prehensile tails, such as the spider monkey, display some homosexual behavior. In the Old World primates, though, homosexual courtship becomes prominent. These primates, including macaques, baboons, gibbons, orangutans, gorillas, chimpanzees, bonobos, and humans, have the most advanced primate societies and the most extensive homosexuality. In these societies, individuals form complex relationships, fostered by both same-sex and other-sex sexual behavior.
When confronted with evidence of homosexuality in nature, most people immediately wonder why two males would waste their time mating with one another rather than mating with females. The answer is that somehow they are not wasting time—that in their present circumstances, the benefits of the male bonding outweighs the benefits of pursuing a heterosexual courtship.
Darwin and Sexual Selection
The diversity of gender and sexuality in nature poses foundational difficulties for biology. Biologists conceptualize gender and sexuality according to Darwin’s The Descent of Man, and Selection in Relation to Sex, which presents the theory of “sexual selection.” Darwin’s theory of natural selection is about evolutionary change in general, whereas the theory of sexual selection is solely about traits pertaining to courtship and mating. The classic example is the peacock’s tail. Darwin assumed that the peahen, the female of the species, was in a position to choose the male she would mate with, and she’s most impressed by male with the showiest plumage. Over many generations, males evolved the showy traits that females prefer.
The issue we face is not whether this story ever occurs, but whether it is representative of how gender and sexuality are expressed throughout nature. That is to say, the preceding examples of animal species that depart from a simple sex binary, that exhibit diverse sex roles, and that express natural homosexuality all refute any claim to generality on the part of sexual-selection theory. The picture of sex and gender expression expressed in Darwin’s theory of sexual selection is not only far from universal, but is perhaps exceedingly rare.
Gender Across Cultures
The comparative anthropology of gender expression across the world and through history is poorly documented relative to that concerning sexuality, though the 1994 classic by Gilbert Herdt, Third Sex, Third Gender: Beyond Sexual Dimorphism in Culture and History, remains timely even today. That said, the available information establishes that transgender-like and multiple gender expressions are vastly more common that we in the West have been led to believe. As recently as fifteen years ago the medical-industrial complex was still floating the figure of roughly one in 10,000 people as transsexual. The latest figures show that around one in 300 is transgender in the U.S.
And as we look beyond the contemporary West, we encounter a huge variety of cultural forms and institutions inhabited by transgender and third gender people. Among the best studied transgender-like cultural forms are the so-called “two-spirit” people of many Native American nations and tribes. The name suggests that they are people who possess a combination of feminine and masculine characteristics.

These people include male-bodied individuals living as women and female-bodied individuals living as men. Here, at left, is a 1900 photograph of Osh-Tish, a well-known male-bodied two-spirit person from the Crow Nation of present-day Wyoming/Dakotas who lived as a woman. At right is an 1890 photograph of a female-bodied two-spirit person from the Quechuan area of Northwestern South America who lived as a man, specifically as a warrior. This warrior, dressed in male clothing, presents in a confident, almost jaunty manner, with no attempt to cover or bind the breasts. The middle panel is a painting of another female-bodied warrior from the Plains Indians. Notice the bare breasts. Two-spirited people assume cross-gender occupations. They do not try to hide or modify their anatomy, because in these cultures occupation rather than genital morphology is the marker of gender identity.
Transgender and gay people are often accused of recruiting young boys into their way of life. Invariably, the truth is that the young people themselves seek out older transgender and gay people, whom they see as kindred spirits. Anthropologist Walter Williams, in his 1986 book The Spirit and the Flesh: Sexual Diversity in American Indian Culture, describes the ceremony by which young boys from several Indian tribes announce their gender and are welcomed by the tribe.
Among the Mohave tribes along the Colorado River in the American Southwest, Williams writes about how parents deal with a boy who has a predisposition to be two-spirited. When the child is about ten years old, his relatives begin discussing his predispositions. The relatives then prepare for a ceremony without letting the boy know about it. The ceremony is meant to take him by surprise, and to serve as both an initiation and a test of his true inclinations. People from various settlements attend. The boy is led into a circle. If he remains there, he accepts that he will go through with the ceremony. A singer sings songs. If the songs move him, he will dance as women do, with intensity. If the boy dances as a woman for four songs, his status as a “two-spirit” is confirmed. Then he is taken by the women, bathed, receives a skirt, returns to the crowd, and announces his new feminine name.
Among the Tohono O’odham Indians of the Sonoran Desert of the American Southwest, the ceremony also involves a public test of inclination. At a social gathering of family and friends, a small brush enclosure is built. A man’s bow and arrows and a woman’s basket are placed inside. The boy is then brought to the enclosure. As the adults watch, he is told to enter the enclosure. The adults set fire to the enclosure. The boy has time to take only one of the two items. If he takes the basket and leaves the bow and arrows, he is confirmed as a two-spirit.
As Williams notes, in all these practices the role of a two-spirit is not forced on the boy by others. While adults might have their suspicions about the boy’s inclinations, it is only when the boy makes the proper move that he is considered “two-spirited.” By doing woman’s dancing, preparing a meal, or taking the woman’s basket, the boy is making an important symbolic gesture. Indians do not see the assumption of two-spirit status as a free choice by the boy but rather as a revelation of his underlying character.
Trans and gay people are too common to represent a pathology. The existence of transgender people throughout the world confirms that trans people comprise a significant proportion of the human species, now and in the past. The reality, extent, and naturalness of trans people do not need to be vouched for by science. Unfortunately, the Western perspective on transgender people relies on a medical construction of transgender experience and identity. That construction presumes a heterosexual gender binary as the standard and seeks to explain variation from that binary as a pathology.
But this medical construction is a fallacy, a scientific mistake that reflects an ignorance of elementary population genetics. For a trait to be considered a pathology, it must be deleterious. It has been known in population genetics for nearly 100 years that there’s a connection between how rare a pathological trait is and how deleterious it is: the more deleterious, the less frequent its occurrence. For example, almost lethal traits like Huntington’s disease are present at five per 100,000 births; while hemophilia A, also rare, occurs at one birth per 8,500.
Gay and transgender people are nowhere close to being this rare. According to the most recent 2011 demographic information from the Williams Institute at UCLA, in the U.S. some 3.5 percent of adults identify as lesbian, gay, or bisexual, and an estimated 0.3 percent of adults identify as transgender. Thus around nine million Americans identify as LGBT, a figure roughly equivalent to the population of the state of New Jersey.
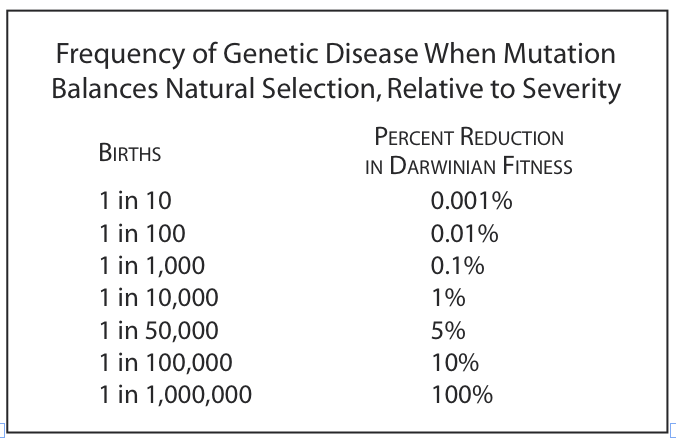 The degree of rarity for pathology is set by a balance between the rate at which the pathology arises by mutation and the rate at which it is eliminated by natural selection. This balance point is called a mutation-selection equilibrium. This table shows the balance point between rarity and deleteriousness, assuming a standard mutation rate of healthy to pathology of one in one million. Rarity is measured in terms of births. Deleteriousness is measured in terms of the percentage loss of survival and/or fecundity caused by the pathology (called “Darwinian fitness” in population-genetic jargon).
The degree of rarity for pathology is set by a balance between the rate at which the pathology arises by mutation and the rate at which it is eliminated by natural selection. This balance point is called a mutation-selection equilibrium. This table shows the balance point between rarity and deleteriousness, assuming a standard mutation rate of healthy to pathology of one in one million. Rarity is measured in terms of births. Deleteriousness is measured in terms of the percentage loss of survival and/or fecundity caused by the pathology (called “Darwinian fitness” in population-genetic jargon).
If the pathology is lethal (bottom line of table), then the trait is exceedingly rare, i.e., one in a million representing a fresh mutation in each instance. For pathologies that are only slightly deleterious, the pathology become much more common, as indicated in the lines toward the top of the table. Deleteriousness of less than, say, one percent is negligible in its impact on evolution. The chance of being killed or failing to reproduce from some random event in life is far higher than that. A small pathology is indistinguishable from the ordinary risks in life.
So, compare the rarity of gay and transgender people with what the table says their deleteriousness would be if they had a pathology. For gays, their rarity lies between the top two lines of the table. For trans people, their rarity lies between second and third lines of the table. For both, the deleteriousness is effectively nonexistent, indistinguishable from the ordinary risks of life, given how common gay and trans people actually are.
Consequently, the scientific validity of the medical construction of how to view people who differ from the heterosexual gender binary must be rejected once and for all. Instead, a future project can be envisioned that seeks to uncover what the positive benefits are to the human variation in gender and sexuality now being mistakenly pathologized in Western culture.
Joan Roughgarden, biology professor emerita at Stanford, is the author of Evolution’s Rainbow; Diversity, Gender, and Sexuality in Nature and People (2004). This piece is excerpted and adapted from an article, “The Gender Binary in Nature, across Human Cultures, and in the Bible,” which appears (in English) in Transsexualität in Theologie und Neurowissenschaften, Gerhard Schreiber, ed. (Walter De Gruyter Inc., 2016).




{kind=link}